| Informatie |
| Home |
| Inleiding |
| Spieren |
| Training |
| Uithoudingsvermogen |
| Rust en herstel |
| Bezoek aan de VU |
| Spieren Elke belasting heeft als eerste effect op de spiercel. De onderdelen van een spiercel hebben andere namen dan de onderdelen van een normale cel. We bekijken een normale cel, en geven daarbij (cursief gedrukt) de namen in een spiercel. Elke cel bevat een celmembraan (sarcolemma) De selectieve permeabiliteit (de doorlaatbaarheid van het celmembraan) voor organische substanties en elektrolyten en de mogelijkheid tot samengaan met andere cellen toont aan dat het celmembraan een zeer ingewikkelde en zeer gespecialiseerde structuur heeft. Het bevat ook alle middelen die nodig zijn voor actief transport, zoals de natriumkalium-pomp tijdens de repolarisatiefase van het celmembraan nadat de actiepotentiaal heeft plaatsgevonden. In het cytoplasma (sarcoplasma), een elektrolyt- en eiwithoudende vloeistof vinden verschillende stofwisselingen plaats. De anaërobe energiestofwisseling (glycolyse), de glycogeensynthese (glycogeen is de intracellulaire opslagvorm van glucose), de glycogeenafbraak en ook de vetzuursynthese vinden plaats in het cytoplasma. In het cytoplasma bevinden zich ook verschillende energieopslagplaatsen, zoals de glycogeenkorrels en de vetdruppeltjes. Het endoplasmatisch reticulum (sarcoplasmatisch reticulum), afgekort ER, strekt zich over het hele cytoplasma uit en vormt het intracellulaire transportsysteem. Op het ER zitten overal ribosomen. Het ER en de ribosomen zorgen samen voor de eiwitsynthese. Het sarcoplasmatisch reticulum heeft in een spiercel een belangrijke functie bij de prikkeloverdracht van het celoppervlak naar het contraktiele fibrillensysteem. De celkern (idem) bevat het genetische materiaal, en speelt ook een belangrijke rol bij de eiwitsynthese. Door de celkern en de ribosomen wordt vermeerdering van eiwitstructuren mogelijk gemaakt. Hierdoor wordt toename van de dikte van een spier mogelijk gemaakt. De mitochondriën (idem) zijn de energiecentrales van een cel. De mitochondriën zorgen voor de oxidatie (=verbranding) van energierijke stoffen. In de mitochondriën bevinden zich de enzymen van de citroenzuurcyclus en de ademhalingsketen. De citroenzuurcyclus is het laatste stadium van de afbraak van koolhydraten, vetten en eiwitten. Al deze stoffen kunnen namelijk, worden afgebroken tot azijnzuur.
In de citroenzuurcyclus wordt er ook weer voor gezorgd dat er weer oxaalazijnzuur wordt opgebouwd, waardoor het proces weer opnieuw kan beginnen. Het oxaalazijnzuur kan zo weer opnieuw een verbinding aangaan met acetyl co-enzym A, waardoor de cyclus weer opnieuw begint. De citroenzuurcyclus levert elke keer weer moleculen ATP. De ademhalingsketen is een systeem van enzymen dat een
onderdeel is van de celademhaling. Dit systeem bevindt zich in het
binnenmembraan van de mitochondriën. Het doel van de ademhalingsketen is alle energie die aanwezig is in een spiercel vast te leggen in het ATP, zodat de spiercel snel over energie kan beschikken. De ademhalingsketen ontvangt de benodigde waterstofatomen van de co-enzymen NAD en FADH2, die deze waterstof in de glycolyse, in de citroenzuurcyclus of tijdens de afbraak van vetzuren en eiwitten hebben overgenomen. De elektronen, die van deze waterstof afkomstig zijn, worden van de ene schakel van de keten naar de volgende overgedragen, totdat de laatste schakel (cytochroom-a/a3) de elektronen overdraagt aan zuurstof. Als daar ook nog waterstofionen aantoegevoegd worden, ontstaat daaruit water (2 e- + 2 H+ + O2 → H2O). De ademhalingsketen bestaat uit drie grote enzymcomplexen die op vaste plaatsen in het binnenmembraan van de mitochondriën liggen. Het eerste complex heeft een lage redoxpotentiaal ( =een hoge elektronendruk), het volgende complex heeft een hogere redoxpotentiaal en het laatste complex heeft de hoogste redoxpotentiaal. Op deze manier kunnen de negatief geladen elektronen van een lage naar een hoge potentiaal worden getransporteerd. Hierbij komt energie vrij. De overdracht van deze negatief geladen elektronen van het ene op het andere complex vindt plaats met behulp van de stoffen ubichinon en cytochroom c. Deze stoffen kunnen zich vrij in het binnenmembraan van de mitochondriën bewegen. De drie enzymcomplexen bestaan uit een combinatie van ijzer-zwavel eiwitten en de katalysator cytochromen (b, c1, a/a3). Beide typen eiwitten bevatten ijzeratomen die bij elektronenoverdracht wisselen tussen de geoxideerde (driewaardige = Fe3+) en gereduceerde (tweewaardige = Fe2+) toestand. Het cytochroom a/a3, dat onderdeel is van het derde complex, dat zuurstof omzet in water, bevat naast ijzer- ook koperatomen die wisselen tussen de één- en de tweewaardige toestand (Cu+, Su2+).
Paragraaf 1.2: Spiervezels
Gladde spieren komen voornamelijk voor in de wanden van onwillekeurig werkende organen zoals bijvoorbeeld het spijsverteringskanaal, luchtwegen, bloedvaten, urinewegen en de baarmoeder. Gladde spieren zijn opgebouwd uit afzonderlijke langwerpige spiercellen en ze staan onder invloed van het autonoom zenuwstelsel, waardoor ze onwillekeurig zijn.
De skeletspier bestaat uit meerdere dwarsgestreepte spiervezels. Elke dwarsgestreepte spiervezel wordt omgeven door een bindweefselschede, het endomysium. Een aantal spiervezels samen vormt een spiervezelbundel die door een bindweefsellaag, het perimysium omgeven wordt. De hele spier bestaande uit heel veel spiervezelbundels wordt omgeven door het epimysium. Spiervezels lopen over het algemeen van pees tot pees. Spiervezels zijn meerdere centimeters lang, hebben een doorsnede van 10 tot 100 µm en bevatten vele langgerekte kernen. De spiervezels worden omgeven door het membraam van de spiercel: het sarcolemma. De spiervezels bestaan zelf uit duizenden vezelvormige elementen, die in de lengterichting van de spier lopen: de myofibrillen. Deze myofibrillen worden door het sarcoplasma met mitochondriën en andere celonderdelen omgeven. De myofibrillen zijn verantwoordelijk voor de contractiele eigenschappen van de spiervezel. In het onderstaande plaatje is te zien hoe een spier is opgebouwd. Een spier bestaat uit een heleboel spiervezelbundels. Die spiervezelbundels bestaan uit spiervezels. De spiervezels zelf zijn weer opgebouwd uit myofibrillen. De myofibrillen zijn opgebouwd uit twee soorten myofilamenten, die zowel chemisch als wat afmetingen betreft, verschillen. Het dwarsgestreepte patroon van de skeletspieren wordt door de regelmatige en gedeeltelijk overlappende rangschikking van deze twee verschillende soorten myofilamenten veroorzaakt. De dikke myosine-filamenten ( 0,01 µm diameter en 1,5 µm lang) lopen evenwijdig op 0.045 µm van elkaar. Tussen de myosinefilamenten liggen, lopend vanaf een verdichting die bekend staat als de Z-streep, dunne actine-filamenten (diameter van 0.005 µm en 1 µm lang, gerekend vanaf de Z-streep). Door deze rangschikking van filamenten neemt men lichtmicroscopisch lichte en donkere banden waar. Zo is een donkere A-band te zien die veroorzaakt wordt door voornamelijk myosinefilamenten. Ook is er een H-band te zien die uitsluitend myosinefilamenten bevat. De M-streep wordt veroorzaakt door verdikkingen in de myosinefilamenten, waarop zich dwarsverbindingen bevinden die de filamenten op afstand van elkaar houden. Het zien van de lichte I-band wordt veroorzaakt door de aanwezigheid van uitsluitend actinefilamenten. Zowel de actinefilamenten als de myosinefilamenten zijn opgebouwd uit ketens van identieke eiwitmoleculen. De actinefilamenten zijn gehecht aan de Z-streep. Het gebied tussen twee Z-strepen wordt een sarcomeer genoemd. Een sacromeer kan worden gezien als het samentrekkende gedeelte van het contractiele apparaat. De werking De overdracht van impulsen van een motorisch axon op de spiervezels gebeurt via een neuro-muscuclaire synaps. De op het presynaptische membraan gevormde actiepotentiaal zorgt ervoor dat acetylcholine wordt uitgestoten in de synapsspleet (synaptic cleft). Hierdoor depolariseert het postsynaptische membraan gedurende 1 a 2 milliseconde. Er ontstaat dan een zogenaamde eindplaatpotentiaal. Het hangt af van de hoeveelheid uitgestoten acetylcholine of de eindplaatpotentiaal het niveau bereikt van een actiepotentiaal. Als het niveau van een actiepotentiaal gehaald wordt, trekken alle spiervezels die zijn aangesloten op hetzelfde motorisch axon samen. De grootte van zo’n groep spiervezels die door een motorisch axon wordt aangestuurd verschilt heel erg, zo kan een motorisch axon 5 spiervezels aansturen ( buitenste oogspieren) en kan een ander motorisch axon bijvoorbeeld honderden spiervezels aansturen.
Als een spier zich samentrekt (spiercontractie), blijft de A-band constant van lengte, terwijl de lengte van de H- en de I-band afneemt. Dit wordt veroorzaakt doordat de actinefilamenten door de myosinemoleculen met zeer kleine stapjes naar de Z-streep toe worden geduwd. Daardoor komen de Z-strepen dichter bij elkaar te liggen, waardoor de sarcomeren dus ook kleiner worden waardoor ook de spiervezel zich verkort. Als alle spiervezels dit doen zal de hele spier zich samentrekken.
Het actine schuift nu 10 nm tussen het myosine. Als alle myosine koppen van de spier gelijktijdig eenmalig zouden roteren, dan zou de spier zich over een afstand van 1% van de spierlengte verkorten. Bij de verkorting van spiervezels treedt ook splitsing van ATP op.
ATP
+ H2O → ADP
+ H3P04 + energie De energie die ontstaat bij deze reactie wordt gebruikt om het dwarsbruggetje (crossbridge) te laten kantelen, zodat de myosine langs het actine-filament beweegt. Dit is het belangrijkste uit de "sliding filament" theorie van Huxley. Elk bruggetje wordt losgemaakt zodra het gevormd is. Dit gebeurt alleen als er voldoende ATP beschikbaar is. Als dit niet het geval is, blijven de bruggetjes op hun plaats. Waardoor de spier stijver wordt. Er zijn ook omstandigheden, waarin een spier krachtig moet samentrekken, dit betekent dat de spierlengte tijdens een contractie soms gehalveerd wordt moet worden. Om dit probleem op te lossen grijpen de snel roterende myosinekoppen een steeds verder weg gelegen actinebindingsplaats. Door deze verschuiving van de twee filamenten verkorten de sarcomeren. Als de sacromeren verkorten trekt de spier samen. Tijdens de repolarisatiefase van de actiepotentiaal neemt de toegenomen diffusie van Ca2+ (en Na+, K+) weer af tot de rustwaarde en zal het tegengesteld gerichte actieve transport van deze ionen de evenwichtssituatie weer doen herstellen. De repolarisatiefase veroorzaakt de ontspanning van de spier. Bij een spier die in rustfase verkeert, bestaat er een blokkade tussen de myosinekop en de bindingsplaats van het actine. Deze blokkade komt tot stand door de aanwezigheid van het tropomyosine-troponine-complex. Deze blokkade kan weer worden opgeheven met behulp van calciumionen.
Paragraaf 1.3: Twee typen spiervezels
Een spiercel of een spiervezel kan uit verschillende vezeltypen bestaan. Er zijn 2 hoofdtypen vezels: ü De witte, dikke en snelle vezel, ook wel FT-vezel genoemd (fast twitch, snel contraherende vezel). Deze vezel is vooral bij snelle, krachtige en intensieve spieracties actief. Deze vezelsoort wordt soms ook wel met type 2 aangeduid. Soms wordt type 2 ook nog onderverdeeld in type 2a,2b en 2c. De 2a vezels maken gebruik van de aërobe en de anaërobe energielevering. Hierdoor zijn deze vezels geschikt voor bijvoorbeeld een sprint. De 2b vezels zijn alleen geschikt voor korte maximale inspanningen. De 2c vezels zijn nog niet gespecialiseerd, ze kunnen nog worden omgebogen naar type 1 of naar type 2a of 2b. ü De rode, dunne en langzame vezel, ook wel ST-vezel genoemd (slow twitch, langzaam contraherende vezel). Deze vezel wordt bij spierarbeid van geringe intensiteit gebruikt. Deze vezelsoort wordt soms ook wel met type 1 aangeduid. Deze 2 vezeltypes verschillen niet alleen in werking, maar ook in stofwisseling. De FT-vezels bezitten veel energieke fosfaten en glycogeen en de bijbehorende uitrusting met enzymen van de anaërobe energiestofwisseling. De ST-vezels bevatten zelf ook veel glycogeen, maar bevatten meer enzymen van de aërobe energiestofwisseling dan de FT-vezels. Bij de ST-vezels is er minder cytoplasma aanwezig en meer mitochondriën. Hierdoor vindt er in de ST-vezels meer activiteit plaats van de enzymen van de citroenzuurcyclus en meer afbraak van vrije vetzuren. In de FT-vezels vindt echter een grotere activiteit plaats in de glycolyse. De samenstelling van de spieren in je lichaam is genetisch vastgelegd. Als je procentueel meer FT-vezels bezit ben je een geboren sprinter. Je bent dan beter geschikt voor korte maximale inspanningen met snelle contracties, dit vind meestal plaats in anaerobe toestand. Als je lichaam meer ST-vezels bezit dan ben je een geboren duursporter. Door training kan je deze aangeboren verhouding van FT –en ST-spiervezels niet veranderen, maar wel de graad van ontwikkeling en daardoor het volume van deze twee verschillende vezeltypes. Om de ST-vezels te trainen moet het zuurstofsysteem getraind worden, wat zorgt voor een toename van het aantal mitochondrien en van het aantal haarvaten in de spieren. Training van ST-vezels zorgt echter niet voor de vergroting van de spieren. Als men de FT-vezels wil trainen moet men vooral de fosfaat poel en het melkzuursysteem trainen. Paragraaf 1.4: ATP
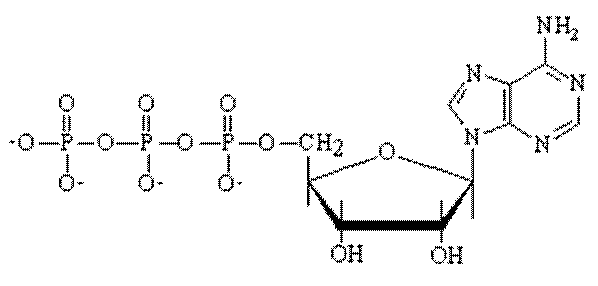
De primaire energiebron voor de spiercontractie is de splitsing van adenosine trifosfaat (ATP). Alle andere energieleverende reacties zoals de aërobe en anaërobe afbraak van koolhydraten en de splitsing van creatinefosfaat zijn geen directe energieleveranciers voor de spiercontractie. Deze andere energieleverend reacties dienen voor de heropbouw van ATP. Adenosine trifosfaat is een relatief klein molecuul, dat wel 4 keer groter is als een glucose molecuul. Het ATP heeft behalve een energieleverende taak ook een 'spier-weekmakende werking'. Zodra de re-synthese van ATP gestopt is, bijvoorbeeld bij een dood iemand, daalt de ATP-spiegel, waardoor de spier verhardt. Een ATP-molecuul bestaat uit 3 delen: Een ringvormig suikermolecuul, een structuur met twee ringen en het belangrijkste deel is het deel met de 3 fosfaatgroepen. De verbindingen tussen de drie fosfaatgroepen zijn energierijk.
Deze bindingen tussen de fosfaatgroepen zijn heel erg zwak. Wanneer de verbinding verbroken wordt komt er veel energie vrij. Bij bijna alles wat je doet heb je ATP nodig. De ATP die nodig is voor de energievoorziening van de cel wordt in eerste instantie verkregen uit de in de cel aanwezige reservevoorraad. De ATP-voorraad in een spiercel bedraagt ongeveer 6 mmol per kg nat spiergewicht en is voldoende voor ongeveer 2 tot 3 seconden spierarbeid. De energie komt op de volgende manier vrij. De buitenste van de drie fosfaatgroepen komt los van het ATP-molecuul. Als deze verbinding is verbroken, ontstaat er ADP (Adenosine difosfaat) en een fosfaatgroep. De fosfaatgroep bevat de meeste energie, deze kan dan weer gebruikt worden om te koppelen aan een ander molecuul. Dit proces wordt fosforylatie genoemd. Als de spierarbeid voorbij is wordt alle ontstane ADP naar het mitchondrium geleid, waar het zich weer koppelt aan een nieuwe fosfaatgroep, waardoor er weer ATP ontstaat. Om de spierarbeid mogelijk te maken nadat de ATP-vooraad op is, moet ATP vrijkomen via verschillende andere wegen. Men maakt daarbij een onderscheid tussen de anaërobe en de aërobe energieleverantie. Het ATP wordt met extreem hoge snelheid door een reactie van ADP met creatine-fosfaat weer aangevuld. Creatine-fosfaat bedraagt ongeveer 20 tot 30 mmol per kg nat spiergewicht en bezit net zoals ATP een energierijke fosfaatgroep. Deze directe vorm van resynthese maakt een totale arbeidstijd (van de gezamenlijke energierijke fosfaten ATP en CP) van maximaal 20 seconden mogelijk. ADP + creatinefosfaat → ATP + creatine De creatine die bij deze ractie vrijkomt wordt voor een deel weer gebruikt voor de resynthese van creatinefosfaat en voor de rest wordt het omgezet in creatinine dat niet verder wordt afgebroken en dat door de nieren wordt uitgescheiden. De energieleverantie in de eerste 7 seconden van de spierarbeid, wordt ook wel de a-lactische fase van de anaërobe energieleverantie genoemd, omdat dit zonder melkzuurvorming plaatsvindt. Aan het begin van iedere belasting met hoge intensiteit, waarbij de energiebehoefte niet voldoende gedekt kan worden, wordt de spier gedwongen de noodzakelijke energie deels langs anaërobe weg te verwerven. Hierbij ontstaat wel melkzuur en wordt daarom de de lactische fase genoemd. Deze fase bevat de (anaërobe) glycolyse. Glucose → enzymen v.d. anaerobe E-leverantie → 2 ATP + melkzuur Deze vorm van energieleverantie vindt plaats in het sarcoplasma bij alle intensieve belastingen waarbij de zuurstofverzorging ontoereikend is. Het maximum van de glycolyse ligt op 45 seconden. Bij de anaërobe glycolyse kan alleen glucose of glycogeen als energiebron dienen. Uit energetisch oogpunt is daarbij het glycogeen dat in de cel aanwezig is gunstiger, omdat het niet eerst via de bloedbaan en door het celmembraan aangevoerd moet worden en dan ook nog is gefosforyleerd moet worden tot ATP. Ook de hoeveelheid ATP die ontstaat bij het gebruik van het glycogeen, dat in de spiercel ligt is hoger. Bij een arbeidsduur die groter is dan een minuut neemt de aërobe energievoorzieningen die in de mitochondriën plaatsvindt in toenemende mate een belangrijke rol in. Bij een oxidatieve verbranding ontstaat: Glucoseà enzymen v.d. aërobe energieleverantieà ATP + CO2 + H2O In tegenstelling tot de anaërobe energievoorziening kan hier naast glucose ook vet en in bijzondere gevallen ook eiwit als brandstof gebruikt worden. De contractiesnelheid van de spiervezel is afhankelijk van de verschillende manieren van energieleverantie. De contractiesnelheid is het hoogst bij het gebruik van energierijke fosfaten en het laagst bij de aërobe verbranding van vetzuren. De verklaring hiervoor ligt in de snelheid waarmee de energierijke fosfaten kunnen worden vrijgemaakt. Als er een hoge intensiteit en daarmee een grote energieomzetting plaats moet vinden, moet er een grotere omzettingssnelheid mogelijk zijn. Is dit niet mogelijk dan leidt dit tot afname van de intensiteit. In het kort wordt ATP met behulp van de volgende stoffen en reacties gemaakt: CP (creatine fosfaat) met ADP, de anaërobe glycolyse en de aërobe energieleverantie. Zij maken ATP vrij, waarbij de afzonderlijke voorraden zich telkens ten koste van de eerstvolgende zullen aanvullen.
|
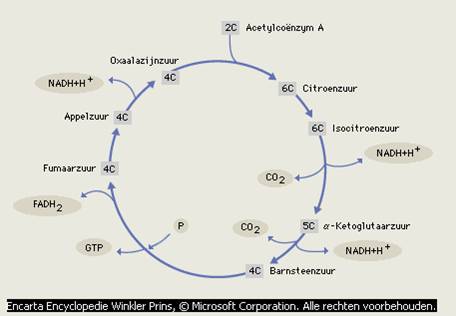 De cyclus begint en
eindigt met een verbinding van acetyl co-enzym A en oxaalazijnzuur die samen
citroenzuur vormen. Deze zuursamenstelling bevat 6 koolstofatomen (4
koolstofatomen van Oxaalazijnzuur en 2 koolstof atomen van acetylcoenzym A ).
Door een reeks van chemische reacties met enzymen tijdens de cyclus worden twee
koolstofatomen afgesplitst.
De cyclus begint en
eindigt met een verbinding van acetyl co-enzym A en oxaalazijnzuur die samen
citroenzuur vormen. Deze zuursamenstelling bevat 6 koolstofatomen (4
koolstofatomen van Oxaalazijnzuur en 2 koolstof atomen van acetylcoenzym A ).
Door een reeks van chemische reacties met enzymen tijdens de cyclus worden twee
koolstofatomen afgesplitst. 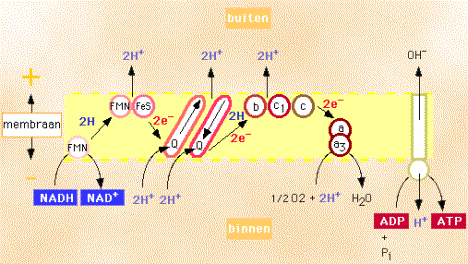
 Er bestaan twee soorten
spieren, die onderscheiden kunnen worden op verschillende eigenschappen van het
weefsel. Het zijn de dwarsgestreepte en de gladde spieren.
Er bestaan twee soorten
spieren, die onderscheiden kunnen worden op verschillende eigenschappen van het
weefsel. Het zijn de dwarsgestreepte en de gladde spieren.  De dwarsgestreepte
spieren zijn te verdelen in twee soorten: de willekeurige dwarsgestreepte
spieren en de onwillekeurig werkende hartspier.
De dwarsgestreepte
spieren zijn te verdelen in twee soorten: de willekeurige dwarsgestreepte
spieren en de onwillekeurig werkende hartspier. Spiercontractie
Spiercontractie 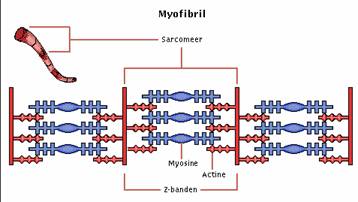 In het sarcoplasma
bevindt zich een systeem van buisjes dat men het sarcoplasmatisch reticulum
noemt. Het bestaat uit twee gedeelten: het transversale systeem (T-systeem) en
het longitudinale systeem (L-systeem). Het T-systeem bestaat uit instulpingen
van het sarcolemma. De inhoud van het T-systeem vormt dus een geheel met de
extracellulaire vloeistof. De T-buisjes komen voor op de grens van de A- en de
I-banden. Het longitudinale systeem (L-systeem) bestaat uit buisjes die
evenwijdig met de myofilamenten meelopen. Ze staan niet in direct contact met
de extracellulaire vloeistof. Het T-systeem en het L-systeem staan in nauw
contact met elkaar. Waar deze 2 systemen contact met elkaar hebben verwijden de
L-buisjes zich tot de laterale cisternen, die samen met het lumen (de holte)
van het T-buisje daar een triade vormen. De T-buisjes zijn verantwoordelijk
voor het snel geleiden van de zenuwprikkel vanaf het sarcolemma naar alle
myofibrillen, zodat daar contractie gelijktijdig kan optreden. Van het
L-systeem
In het sarcoplasma
bevindt zich een systeem van buisjes dat men het sarcoplasmatisch reticulum
noemt. Het bestaat uit twee gedeelten: het transversale systeem (T-systeem) en
het longitudinale systeem (L-systeem). Het T-systeem bestaat uit instulpingen
van het sarcolemma. De inhoud van het T-systeem vormt dus een geheel met de
extracellulaire vloeistof. De T-buisjes komen voor op de grens van de A- en de
I-banden. Het longitudinale systeem (L-systeem) bestaat uit buisjes die
evenwijdig met de myofilamenten meelopen. Ze staan niet in direct contact met
de extracellulaire vloeistof. Het T-systeem en het L-systeem staan in nauw
contact met elkaar. Waar deze 2 systemen contact met elkaar hebben verwijden de
L-buisjes zich tot de laterale cisternen, die samen met het lumen (de holte)
van het T-buisje daar een triade vormen. De T-buisjes zijn verantwoordelijk
voor het snel geleiden van de zenuwprikkel vanaf het sarcolemma naar alle
myofibrillen, zodat daar contractie gelijktijdig kan optreden. Van het
L-systeem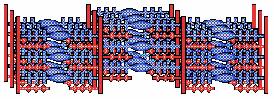 Bij een spier die actief
wordt, veroorzaakt de spieractiepotentiaal een sterke toename van calciumionen
rond de crossbridge. Dit gebeurt op de volgende twee manieren: meer calciumionen
zullen de cel in diffunderen en er volgt een sterke toename van de calciumionen
doordat er ionen uit opslagplaatsen in de cellen vrij komen. De calciumionen
binden zich aan troponine, waardoor de vorm van troponine verandert en waardoor
tropomyosine zijn remmende werking verliest. Tropomyosine verplaatst zich nu in
de diepe groeve van het actinefilament. Op deze manier wordt er een bepaalde
plaats blootgelegd van het actinemolecuul, zodat de myosine kopjes contact
kunnen maken met de actine-moleculen. Er ontstaan dan een soort van bruggen
tussen de actine en de myosine, crossbridges genoemd. Nu wordt de myosinekop
dus steeds volgens dit mechanisme
Bij een spier die actief
wordt, veroorzaakt de spieractiepotentiaal een sterke toename van calciumionen
rond de crossbridge. Dit gebeurt op de volgende twee manieren: meer calciumionen
zullen de cel in diffunderen en er volgt een sterke toename van de calciumionen
doordat er ionen uit opslagplaatsen in de cellen vrij komen. De calciumionen
binden zich aan troponine, waardoor de vorm van troponine verandert en waardoor
tropomyosine zijn remmende werking verliest. Tropomyosine verplaatst zich nu in
de diepe groeve van het actinefilament. Op deze manier wordt er een bepaalde
plaats blootgelegd van het actinemolecuul, zodat de myosine kopjes contact
kunnen maken met de actine-moleculen. Er ontstaan dan een soort van bruggen
tussen de actine en de myosine, crossbridges genoemd. Nu wordt de myosinekop
dus steeds volgens dit mechanisme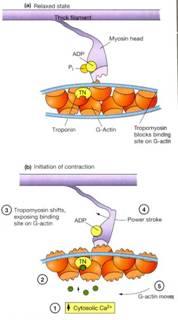 De top van elk myosine
kopje (myosin head) bindt zich aan een
De top van elk myosine
kopje (myosin head) bindt zich aan een